
Dominância de carbono orgânico particulado produzido in situ em um reservatório subtropical inferido a partir de isótopos estáveis de carbono
Este trabalho é somente para uso privado de atividades de pesquisa e ensino. Não é autorizada sua reprodução para quaisquer fins lucrativos. Esta reserva de direitos abrange a totalidade dos dados do documento bem como seu conteúdo. Na utilização ou citação de partes do documento é obrigatório mencionar o nome da pessoa autora do trabalho.
Dominance of in situ produced particulate organic carbon in a subtropical reservoir inferred from carbon stable isotopes
RESUMO
Fontes de carbono orgânico particulado (COP) desempenham papéis importantes no ciclo do carbono aquoso porque a produção interna pode fornecer material lábil que pode ser facilmente transformado em CO2. Por outro lado, entradas externas de COP mais recalcitrantes podem causar aumento de cargas na matéria orgânica sedimentar que pode, em última análise, causar liberação de CH4.
Para diferenciar as fontes, os isótopos estáveis oferecem uma ferramenta útil. Apresentamos um estudo sobre o reservatório de Itupararanga (Brasil), onde as origens do COP foram exploradas comparando suas razões isotópicas (δ13C POC) com aquelas do carbono inorgânico dissolvido (δ13CDIC ).
O δ13C POC teve uma média de cerca de -25,1‰ em águas próximas à superfície, o que indica maior produção primária inferida a partir de um modelo de fracionamento que leva em consideração a transferência de carbono com uma avaliação combinada de δ13C POC , δ13C DIC e CO2 aquoso. No entanto, os valores de δ 13 C POC para profundidades de água de 3 a 15 m diminuíram para -35,6‰ e indicaram diferentes fontes de carbono. Consequentemente, os valores de δ 13 CDIC do reservatório tiveram uma média de cerca de +0,6‰ nos 3 m superiores da coluna d’água. Isso indica desgaseificação de CO2 e fotossíntese.
Abaixo dessa profundidade, valores de isótopos de DIC tão baixos quanto -10,1‰ mostraram influências mais fortes da respiração. Um modelo de fracionamento com ambos os parâmetros isotópicos revelou que 24% do POC no reservatório se originou de detritos fora do reservatório e 76% dele foi produzido internamente pela fixação de CO2 aquoso.
Palavras-chave: carbono orgânico, isótopos, fotossíntese, COP
ABSTRACT
Sources of particulate organic carbon (POC) play important roles in aqueous carbon cycling because internal production can provide labile material that can easily be turned into CO2. On the other hand, more recalcitrant external POC inputs can cause increased loads to sedimentary organic matter that may ultimately cause CH4 release.
In order to differentiate sources, stable isotopes offer a useful tool. We present a study on the Itupararanga Reservoir (Brazil) where origins of POC were explored by comparing its isotope ratios (δ13CPOC) to those of dissolved inorganic carbon (δ13CDIC). The δ13CPOC averaged around − 25.1‰ in near-surface waters, which indicates higher primary production inferred from a fractionation model that takes into account carbon transfer with a combined evaluation of δ13CPOC, δ13CDIC and aqueous CO2. However, δ13CPOC values for water depths from 3 to 15 m decreased to − 35.6‰ and indicated different carbon sources.
Accordingly, the δ13CDIC values of the reservoir averaged around + 0.6‰ in the top 3 m of the water column. This indicates CO2 degassing and photosynthesis. Below this depth, DIC isotope values of as low as − 10.1‰ showed stronger influences of respiration. A fractionation model with both isotope parameters revealed that 24% of the POC in the reservoir originated from detritus outside the reservoir and 76% of it was produced internally by aqueous CO2 fixation.
Keywords: organic carbon, isotopes, photosynthesis, COP
Introdução
Reservatórios e seus afluentes são componentes ativos das paisagens. Eles recebem, transportam, processam e armazenam carbono inorgânico e orgânico 11 . Embora lagos e reservatórios cubram apenas 2,2% da área continental global, eles podem desempenhar papéis até então pouco contabilizados na ciclagem do carbono continental 2 , 3 , 4 . Além disso, investigações regionais de orçamentos de carbono tornam-se cada vez mais importantes para a gestão dos recursos hídricos, pois definem funções e serviços ecossistêmicos 5 , 6 , 7 , 8 .
Mudanças espaço-temporais de parâmetros físico-químicos e biológicos em reservatórios subtropicais podem aumentar nosso conhecimento sobre a rotatividade de carbono em corpos d’água. Por exemplo, precipitação excessiva pode aumentar nutrientes terrestres, contaminantes, lixiviação do solo e adições de entrada de esgoto não tratado 9 . Eventos de chuva também podem causar turbulências próximas à superfície em lagos, o que demonstrou aumentar a troca gasosa aumentando as velocidades de transferência de CO 2 10 , 11 .
A precipitação também pode levar ao aumento da entrada de fontes externas (ou seja, matéria alóctone, por exemplo, de restos de plantas ou solos) que podem ficar presos em lagos e reservatórios. Isso pode, por sua vez, alterar a dinâmica biogeoquímica da coluna d’água 12 . Tais processos incluem a absorção de carbono e a liberação de carbono dentro da coluna d’água, que também podem ser controlados por mudanças sazonais de temperatura, padrões climáticos ou disponibilidade de luz. Estes podem, por sua vez, ter influências importantes nos fluxos de carbono dentro de corpos de água abertos, como por exemplo sedimentação, ou produção primária bruta, respiração do ecossistema e entradas externas de carbono 13 .
Para este último, as bacias hidrográficas dos reservatórios podem oferecer contribuições grandes, mas desconhecidas, que podem exceder a produção primária interna (ou seja, autóctone) 14 . Por outro lado, o POC autóctone e a mineralização de sedimentos são fortemente restringidos 15 e o excesso deste tipo de carbono orgânico pode aumentar a emissão de gases de efeito estufa em 20–70% (de acordo com a taxa de mineralização), principalmente durante períodos de eutrofização 16 .
Devido ao importante papel do POC como fonte ou sumidouro de carbono, uma tarefa essencial é determinar a origem dessa fração de carbono em ecossistemas aquáticos. Por exemplo, mudanças nas fontes terrestres e lacustres de POC pela fotossíntese podem afetar o ciclo interno do carbono orgânico (CO) na coluna d’água 17 , 18 , 19 , 20. Um melhor conhecimento dessas fontes e sumidouros de carbono também pode ajudar a estabelecer importantes ferramentas de decisão para a gestão da qualidade da água. Isso inclui a prevenção e o controle da eutrofização e da poluição, da degradação de bacias hidrográficas e de processos relacionados à paisagem, incluindo transporte, transformação e deposição de material terrestre em ecossistemas aquáticos 21 , 22 .
A vida aquática depende de fontes de carbono alóctones de detritos terrestres e solos, mas também pode depender de fontes autóctones, como macrófitas e algas. Os padrões de processamento dessas fontes de carbono orgânico são diferentes. Em geral, o CO alóctone é mais recalcitrante e tem taxas de degradação mais baixas e, portanto, muitas vezes se torna preferencialmente sequestrado em sedimentos 23 , 24 , 25 , 26 , 27.
Em contraste, o CO autóctone da produção primária geralmente sofre rápida renovação 24. Isso mostra a importância de entender as extensões desses membros finais 28 , 29. Isso também se reflete na literatura científica recente que tem demonstrado crescente interesse na dinâmica do POC, especialmente por meio de isótopos estáveis de frações de carbono 21 , 30 , 31 , 32 , 33 .
Essa demanda por melhor compreensão da dinâmica do POC também se relaciona à importância de ferramentas integrais para estudar a produção e o consumo de carbono inorgânico dissolvido (DIC) e CO 2 por meio da decomposição microbiana, desgaseificação, fotossíntese e respiração em sistemas aquáticos 17 , 34 , 35 , 36 , 37 . Tais abordagens dificilmente foram aplicadas a reservatórios subtropicais (e até agora em nenhum lugar no Brasil), onde o processamento de carbono pode ser particularmente intenso devido às temperaturas quentes.
Este estudo de caso foi realizado no reservatório de Itupararanga, um importante corpo de água doce no estado de São Paulo (Brasil) e monitorado desde 1998 pela Companhia Ambiental do Estado de São Paulo 38 [ https://cetesb.sp.gov.br/aguas-interiores/publicacoes-e-relatorios/ ]. Este monitoramento revelou que este reservatório subtropical enfrenta deterioração progressiva da qualidade da água nas últimas três décadas (Fig. 1 ). Tais processos e o ciclo de carbono relacionado foram investigados pelas razões POC/chl-a e também pelas razões combinadas de isótopos de carbono de aplicação (δ 13 C POC e δ 13 C DIC ) e conteúdos aquosos de CO2 .
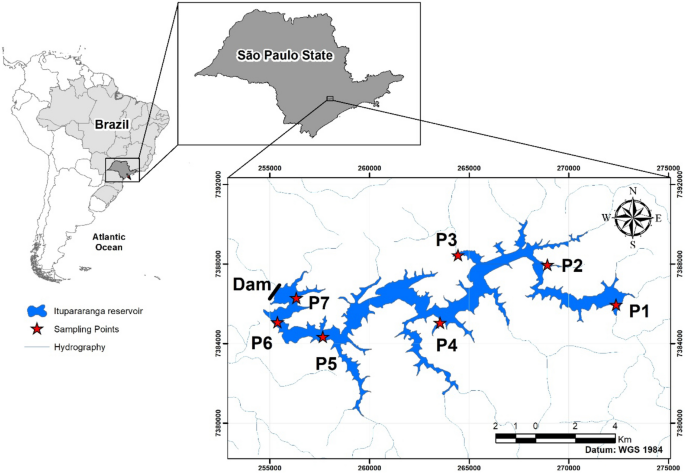
Este reservatório subtropical é econômica e ambientalmente importante para a região metropolitana de São Paulo. A deterioração geral da qualidade ambiental e da água do sistema Itupararanga atraiu o interesse de vários pesquisadores 39 , 40 , 41 , 42 , 43 . Esses estudos consideraram aspectos de nutrientes e comunidades de cianobactérias, macroinvertebrados sedimentares, toxicidade de metais e uso da terra em relação à qualidade da água. Este trabalho mostrou que o reservatório de Itupararanga recebe quantidades substanciais de nutrientes, principalmente devido ao uso de terras agrícolas na bacia hidrográfica circundante 42 .
Outra fonte importante de nutrientes é a descarga de esgoto de áreas urbanas. Eles chegam principalmente ao reservatório sem tratamento na entrada 39 , 41 . Esses insumos podem estimular a produção primária e também podem causar eutrofização. Essa entrada de nutrientes terrestres também sugere que matéria orgânica (MO) e material detrítico podem entrar no reservatório por córregos e escoamento superficial através de uma extensa linha costeira. No entanto, menos atenção tem sido dada a outras fontes de carbono neste sistema.
Portanto, o objetivo deste estudo é avançar o conhecimento sobre a origem interna do reservatório de POC, explorando variações espaço-temporais dessa fração de carbono, juntamente com as razões de isótopos estáveis de DIC e POC. Isso também ajuda a delinear os aportes terrestres. Essa técnica, até o momento, raramente foi aplicada ao reservatório de Itupararanga, que pode ser considerado um corpo d’água subtropical, e apresentamos a abordagem como uma ferramenta para compreender melhor os sistemas hídricos de ciclagem de carbono.
Resultados
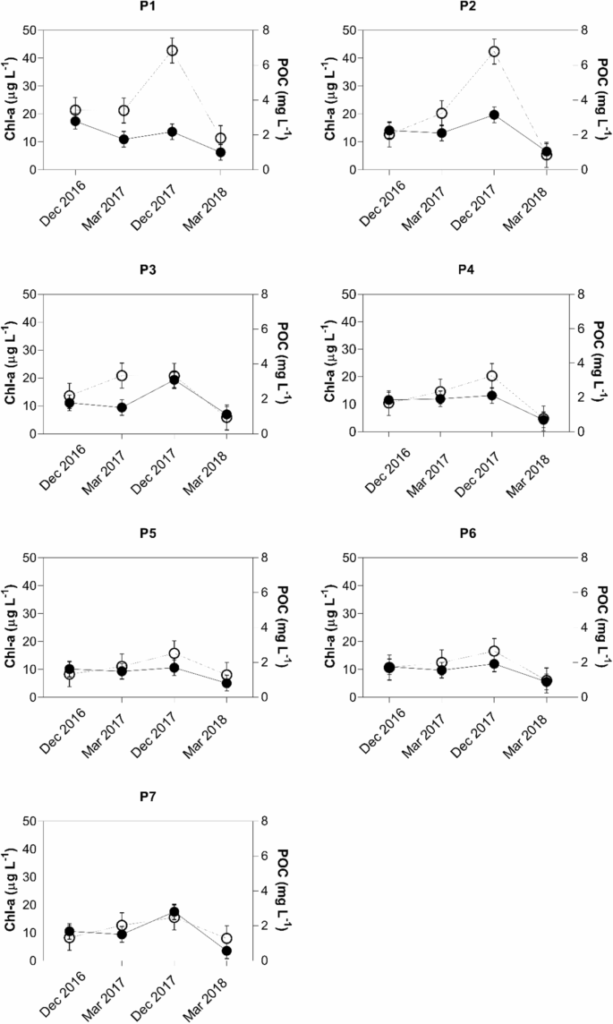
Concentrações aumentadas de POC e chl-a foram observadas em todos os locais de amostragem de superfície em profundidades acima de 3 m em dezembro de 2017, com valores máximos de 2,2 mg L −1 e 42,8 μg L −1 para POC e chl-a (Fig. 2 ). Após esta temporada, uma diminuição das concentrações de chl-a e POC foi encontrada em todos os sete locais de amostragem entre dezembro de 2017 e março de 2018.
A diminuição média foi de 67% para chl-a e 62% para POC e uma diminuição máxima de chl-a de 87% foi encontrada em P2. A diminuição máxima para POC foi de 80% em P7. Os valores de POC e chl-a nos sete locais de amostragem no reservatório de Itupararanga são exibidos na Fig. 2 .
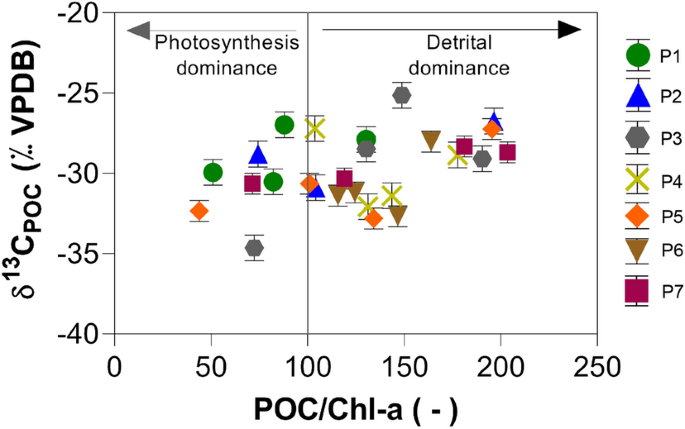
Em uma primeira tentativa de delinear as fontes, plotamos as razões POC/chl-a em relação aos valores de (Fig. 3 ). Aqui, usamos um limite de 100 para as razões POC/chl-a para definir a dominância da fotossíntese sobre a entrada detrítica 44 . De acordo com essa abordagem, a produção primária domina em apenas 26% das amostras de água superficial. Essas amostras são de P1 em março de 2017, dezembro de 2017 e março de 2018, de P2 em dezembro de 2017, de P3 em março de 2017, de P5 em dezembro de 2017 e de P7 em março de 2018.δ
Para testar como o carbono pode ser trocado entre DIC e POC, investigamos seus valores de isótopos estáveis em uma correlação de Spearman. A correlação indicou tendências positivas e significativas com valores de r entre 0,79 e 0,98 e valores de p entre menos de 0,0001 e 0,0182 (Tabela 1 ).Tabela 1 Coeficientes de correlação de Spearman ( r ), valores de p ( p ) e número de amostras (n) para versus .δδ
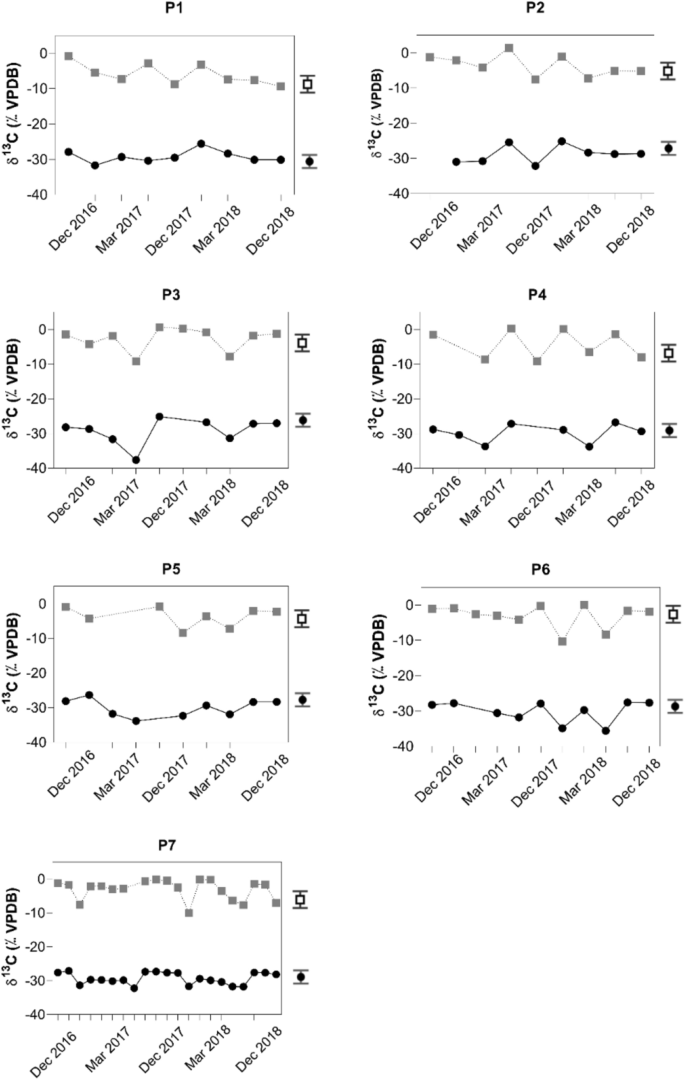
Amostras de profundidades abaixo de 3 m apresentaram valores mais baixos para que variaram de —10,4‰ a —7,0‰ para todas as campanhas. Os valores representativos deste grupo são —8,8‰ para P1 em dezembro de 2017 a 3 m, —7,5‰ para P2 em dezembro de 2017 a 5 m, —9,2‰ para P3 em março de 2017 a 11 m, —9,2‰ para P4 em dezembro de 2017 a 8 m, —8,4‰ para P5 em dezembro de 2017 a 11 m, —10,4‰ para P6 em dezembro de 2017 a 15 m e —10,1‰ para P7 em dezembro de 2017 a 14 m de profundidade (Fig. 4 ).
Para os mesmos locais de amostragem, amostras representativas da superfície apresentaram mais positivos com + 1,5‰ para P2 em dezembro de 2017 a 1 m, + 0,6‰ para P3 em dezembro de 2016 a 1 m, + 0,3‰ para P4 em dezembro de 2017 a 1 m e + 0,04‰ para P6 em março de 2018 a 1 m de profundidade (Fig. 4 ).δ{\updelta }^{13} {\texto{C}}_{{{\texto{DIC}}}}δ
Fracionamento de isótopos de carbono
Os dados também foram plotados de acordo com um modelo de fracionamento de Rau et al. 55 . Nessa abordagem, os conteúdos aquosos de CO 2 são plotados em função das diferenças isotópicas entre e de acordo com as Eqs. ( 2 )–( 5 ) (Fig. 5 ). O mesmo foi repetido para amostras de superfície a fim de proporcionar a melhor comparação com os dados da razão POC/chl-a (Fig. S1 ).δδ
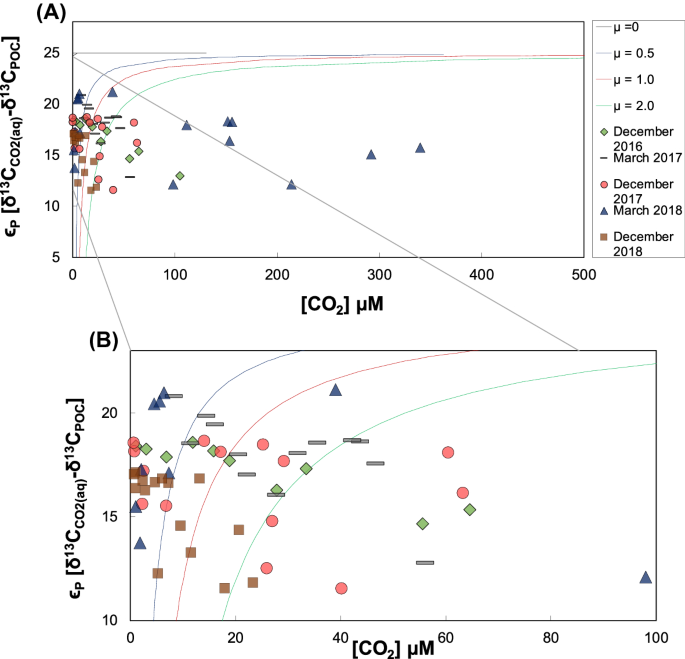
A Figura 5 mostra que 80% das amostras coletadas durante o período de dezembro de 2016 a dezembro de 2018 concordam com o modelo de fracionamento por produtores primários. Em contraste, o sinal de POC possivelmente detrítico (amostras que não se encaixaram no modelo de fracionamento Rau) apareceu nas estações quentes de dezembro de 2017 e março de 2018, respectivamente (Tabela S10 — material suplementar). Ao tratar apenas as amostras de superfície com esta abordagem, 95% de todas as amostras se ajustaram ao modelo (Fig S1 ). As que não concordaram com o modelo de fracionamento são amostras de P1 em dezembro de 2017 e março de 2018, ambas a uma profundidade de água de 3 m.
Discussão
The possible significance of subtropical reservoirs in global and regional carbon budgets is still vague. One reason for this is the complexity and variability of external carbon inputs to these systems. Here, we investigated seasonal and spatial variations of POC and chl-a. These parameters can be combined to POC/chl-a ratios. However, this approach only offers a subjective indicator for dominance of external or internal POC sources. Combined investigation of δ13CPOC and δ13CDIC values seem more promising to help separating POC sources, because this technique also maps transformations of carbon. Therefore, the application of a carbon stable isotope fractionation model should be able to provide distinct information on POC sources.
When looking at concentrations of POC and chl-a, most samples in Fig. 2 showed similar spatio-temporal distributions of both parameters. Note that P1 showed no considerable changes in chl-a in December 2016 and March 2017 (i.e. the warm season with precipitation of 148 and 146 mm, respectively).
This observation was made even though the corresponding POC concentrations decreased. However, March 2017 and December 2017 had similar precipitation patterns with averages of 148 and 157 mm, respectively, but both chl-a and POC increased. Despite similarities of rainfall averages in both sampling periods, individual intense events may have increased the flushing of nutrients into the reservoir. This may in turn have caused POC generation by primary production. These trends agree with the findings of other studies44,45,46.
Sampling locations P2, P3, P5, P6 and P7 showed decreasing trends in POC from December 2016 to March 2017. These were not matched by increasing chl-a concentrations until the end of December 2017. This trend might be related to declining inputs of detrital material, organic matter consumption, or sedimentary deposition. On the other hand, increases in chl-a contents in the same period was likely caused by supply of phytoplanktonic POC. This was confirmed by an observed algae bloom in the same period, according to the Inland Water Quality Report, produced by CETESB (São Paulo State Environmental Company) in 2017. Higher concentrations of both parameters shown in Fig. 2 may also reflect algae blooms in the reservoir in December 2017.
Valores baixos de POC/chl-a na Fig. 3 indicaram uma dominância da fotossíntese (fitoplâncton) no pool de POC. Observações semelhantes também foram feitas por outros autores em outros sistemas aquosos abertos 44 , 47 . Embora não seja possível detectar padrões sazonais e espaciais claros a partir dessa abordagem, mais da metade das amostras foram classificadas com origem de dominância detrítica (Fig. 3 ). O outro grupo de amostras pertence aos períodos quentes de março de 2017, dezembro de 2017 e março de 2018. Todas parecem ser caracterizadas por uma dominância da fotossíntese. No entanto, alguns pontos amostrais desses eventos de amostragem também foram plotados no campo detrítico da Fig. 3 .
Embora todas as amostras para as medições de chl-a pertençam à superfície ( 3 m de profundidade), suas razões POC/chl-a sugerem que 74% do POC no reservatório de Itupararanga se originam de detritos fora do reservatório. Uma explicação é que a razão limite POC/chl-a de 100 pode servir apenas como um indicador aproximado e arbitrário para a entrada de carbono de algas versus detrítico.
Além disso, esse valor limite pode variar ao longo do tempo e do espaço. No geral, seria de se esperar que muito mais amostras fossem plotadas no campo da fotossíntese ao considerar as entradas de nutrientes conhecidas no reservatório e as florações de algas observadas. Uma conclusão intermediária é que o pool de POC é mais complexo e não pode ser explicado apenas com dados de concentração de chl-a e POC. Isso seria plausível, porque a produção primária em comunidades complexas impacta os ambientes aquáticos por meio de inúmeras interações metabólicas 48 .
A correlação de Spearman na Tabela 1 indica fortes relações entre e . Este é um bom indicador da possibilidade de que ambas as variáveis possam ter sido afetadas por processos semelhantes, sendo a produtividade biológica a mais plausível. Isso significa que os processos que afetam o fracionamento de isótopos de DIC resultam da transformação desta fonte de carbono por meio da produção (fotossíntese) e utilização (respiração) de carbono orgânico.δδ
Mais valores enriquecidos em 12 C de variaram de —10,4‰ a —7,0‰ e sugerem sinais respiratórios que foram mais óbvios no local de amostragem P1 (Fig. 4 ). Tais valores enriquecidos em 12 C de também foram observados em outros estudos 49 , 50 . O fato de que esses valores foram observados apenas para amostras de profundidades de água de três metros e abaixo é plausível, porque além dessa profundidade a penetração limitada de luz dificulta a fotossíntese. Esse processo geralmente enriquece o DIC restante em 13 C.
Além disso, o aumento da precipitação também pode ter liberado o DIC e o POC empobrecidos em 13 C da bacia hidrográfica circundante 17 . Os valores mais positivos de δ 13 C DIC observados perto da superfície da água provavelmente resultam do equilíbrio com o CO 2 atmosférico e da fotossíntese, que enriquecem o DIC restante em 13 C.δ{\updelta }^{13} {\texto{C}}_{{{\texto{DIC}}}}δ
Para uma investigação mais detalhada, plotamos os dados de acordo com um modelo descrito por Rau et al. 55 . No geral, a maioria das amostras coletadas entre dezembro de 2016 e dezembro de 2018 revelou uma dominância de POC produzido internamente no reservatório. Isso pode indicar um padrão típico para reservatórios tropicais e subtropicais com temperaturas mais altas, aportes críticos de nutrientes e atividades biológicas geralmente aprimoradas. O grupo amostral que está de acordo com o modelo de Rau et al. 55 abrange todas as profundidades de amostragem em todas as estações.
Esta é uma boa indicação de que o POC foi predominantemente produzido de forma autóctone com baixa variabilidade espaço-temporal. Quando o POC fotossintético foi encontrado fora da zona fótica, provavelmente foi produzido próximo à superfície e depois movido para baixo através da coluna d’água. As amostras que não se encaixam neste modelo (Fig. 5 B) indicam uma dominância de POC alóctone que não segue os mesmos padrões de transferência de carbono entre o CO2 aquático e o POC. Isso confirma outros estudos que também mostraram que entradas alóctones produzem diferenças isotópicas entre POC e DIC (ε) que estão abaixo das linhas modeladas por Rau et al. 44 , 51 .
Para uma melhor comparação com o método alternativo de proporções POC/chl-a, consideramos apenas amostras de até 3 m de profundidade de água no Modelo Rau (Fig. S1 em material suplementar). Isso mostra que apenas 5% do subconjunto de dados não concordava com o modelo (ou seja, P1 em dezembro de 2017 e P1 em março de 2018, ambos a 3 m de profundidade). A concordância com o modelo do restante das amostras de água superficial na zona fotossinteticamente ativa é um bom indicador da geração biológica in situ de POC pela fotossíntese.
Essa interpretação do modelo contrasta com os resultados das proporções POC/chl-a (Fig. 3 ), que revelaram contribuições diferentes para fontes autóctones e alóctones de carbono com 26 e 74%, respectivamente. Essa diferença é surpreendente, porque as amostras para chl-a também foram coletadas na zona fótica dos 3 m superiores da coluna d’água. Uma razão para essa discrepância pode ser que mais POC detrítico ocorra no reservatório após fortes chuvas e tempestades. Isso também poderia mobilizar chl-a da paisagem a partir de resíduos vegetais.
Nesse processo, as relações POC/chl-a poderiam ser diferentes daquelas do material de algas recém-produzido e mascarar os efeitos da fotossíntese in situ. Efeitos de mascaramento semelhantes podem existir após a ressuspensão do POC sedimentar. No entanto, a definição arbitrária de 100 das relações POC/chl-a para a separação da fotossíntese e dos detritos é provavelmente a razão mais forte para a discordância entre ambas as abordagens. Por exemplo, definir esse limite para 150 resultaria em cerca de metade das amostras resultantes de entrada terrestre.
O fato de muitos dados se ajustarem ao modelo em diferentes taxas de crescimento específicas (μ) também pode indicar uma sequência sazonal de tipos de produtores primários, como Bacillariophyceae , Achnanthidium minutissimum , F. delicatissima var. delicatissima . Essas variedades de diferentes algas também foram observadas em estudos anteriores 52 , 53 . Essa observação também é apoiada por padrões sazonais de δ 13 C DIC e δ 13 C POC no conjunto de dados. Como os intervalos de sazonalidade investigados abrangem vários meses, o POC autóctone pode ser produzido continuamente em águas superficiais e provavelmente tem efeitos mais persistentes no ecossistema do reservatório do que o POC terrestre 34 .
No geral, considerações simultâneas de δ 13 C DIC e δ 13 C POC , juntamente com a disponibilidade de CO 2(aq) no Modelo Rau, marcam a transferência de carbono da fase DIC para a fase POC. Portanto, deve ser um melhor indicador do uso biológico de carbono na forma de crescimento de algas. Também mostra como o fracionamento de isótopos estáveis de carbono depende de fontes limitantes de taxa e do tamanho da célula. Assim, essa abordagem parece melhor em capturar complexidades de transferência de carbono pelo crescimento de algas 54 .
No entanto, o Modelo Rau só pode prever a assimilação de CO 2 aquoso e não leva em conta a absorção direta de HCO 3 – aquoso da coluna de água. Isso pode ignorar uma parte significativa de uma comunidade de algas que opera dessa maneira 55 . Além disso, em locais próximos à costa, macrófitas ativas podem alterar os resultados influenciando o δ 13 C DIC , que também serve como um parâmetro de entrada para o Modelo Rau. Mesmo que as macrófitas contribuam para o pool de POC por meio de fragmentos de plantas, elas podem seguir diferentes padrões de enriquecimento de isótopos que não foram investigados aqui.
O modelo de fracionamento de Rau et al. 55 foi testado para todos os dados e todos os períodos de amostragem. Ele reduziu com sucesso a complexidade da assimilação de carbono para fracionamento específico, taxas de crescimento (μ) e concentrações extracelulares de CO2 (aq) . Como revelou que a maioria das amostras de lâmina d’água inferior a 3 m (ou seja, a zona fótica) concordava com este modelo, a fotossíntese e o consumo de CO2 (aq) parecem ser os processos dominantes para a produção de POC. Além disso, para algumas amostras coletadas em lâminas d’água superiores a 3 m, o modelo produziu resultados plausíveis, indicando, assim, o transporte vertical de POC produzido biologicamente na coluna d’água.
Ao considerar todos os pontos de dados deste estudo, cerca de vinte por cento das amostras de POC não se encaixaram no modelo e foram plotadas abaixo das linhas teóricas. Essas amostras provavelmente correspondem à entrada detrítica ou à ressuspensão de matéria sedimentar. Esta última pode se originar da ressurgência durante tempestades que, além disso, também pode importar mais POC de fora do reservatório.
Essas descobertas também podem se estender a outros sistemas de água doce e, portanto, oferecem uma ferramenta para separar as vias de entrada de POC que, por sua vez, podem ter controles importantes na ciclagem de CO2 (aq) . Este seria o caso, por exemplo, se mais POC autóctone estivesse presente. Por outro lado, o POC predominante de fontes terrestres pode implicar maior geração de metano a partir de sedimentos. Isso ocorre porque o material alóctone mais recalcitrante geralmente é menos facilmente consumido na coluna d’água e se torna mais provável de ser depositado em sedimentos. Aqui, condições anóxicas podem produzir metano por períodos de tempo mais longos do que os tempos de residência na coluna d’água.
No geral, nossos resultados sugerem que isótopos estáveis podem ajudar a rastrear fontes de POC. Tais aplicações combinadas de isótopos estáveis de carbono e parâmetros físico-químicos são promissoras para fornecer diferenciações sensíveis de fontes de carbono em águas naturais.
Métodos
Local de estudo
O reservatório de Itupararanga está localizado em um clima subtropical que pertence ao tipo Cwb, de acordo com a classificação de Köppen 56 . Está situado na bacia hidrográfica do alto rio Sorocaba, no estado de São Paulo, Brasil (Fig. 1 ). O clima é caracterizado por uma estação seca de abril a setembro e uma estação chuvosa de outubro a março. A temperatura média mensal é superior a 18 °C em todos os meses e atinge 22 °C em dezembro 57.
A Tabela S8 (material suplementar) mostra dados mensais de precipitação de 2016 a 2018 de várias estações meteorológicas próximas ao reservatório de Itupararanga do Instituto Nacional de Meteorologia (INMET) e do Departamento de Águas e Energia (DAAE). O reservatório foi construído em 1912 com o propósito original de geração de energia hidrelétrica. Atualmente, é usado predominantemente como fonte de água potável para aproximadamente um milhão de pessoas, bem como para irrigação e lazer 58 .
O reservatório possui profundidades máxima e média de 21 e 7,8 m, respectivamente. O canal principal tem cerca de 26 km de extensão. O tempo de residência da água varia entre 95 e 270 dias, e o reservatório possui um volume máximo de armazenamento de 286 milhões de m³ 41 , 58. Os córregos contribuintes mais importantes são os rios Sorocabaçu, Sorocamirim e Una, além de inúmeros pequenos córregos (por exemplo, Paruru, Ressaca e Campo Verde) 59 .
Sete locais (P1–P7) dentro do reservatório de Itupararanga foram amostrados entre dezembro de 2016 e dezembro de 2018, em profundidades entre 0 e 15 m abaixo da superfície da água (Fig. 1 ). Dependendo da profundidade da coluna d’água, os intervalos amostrados variaram entre 1 e 5 m (ver Tabelas S1 – S7 no material suplementar).
O uso do solo nas proximidades dos locais de amostragem P1 e P2 é dominado por gramíneas e pastagens, porém P1 também recebe insumos de esgoto. Os locais de amostragem P3 e P4 estão próximos de áreas urbanas, enquanto P5 a P7 estão localizados em partes da bacia hidrográfica dominadas por florestas, agricultura e silvicultura 42 .
Procedimentos no local e laboratoriais
Os perfis de temperatura e pH foram medidos em todos os locais de amostragem utilizando uma sonda multiparâmetro portátil (Horiba U-50) 42 . As amostras para medições isotópicas foram filtradas por filtros de disco de náilon com porosidade de 0,45 μm em frascos de vidro âmbar de 40 mL, de acordo com os padrões da Agência de Proteção Ambiental dos EUA (EPA). Todos os frascos de amostra para medições isotópicas foram preservados com 0,05 mL de uma solução saturada de cloreto de mercúrio (HgCl2 ) para evitar atividades biológicas secundárias após a amostragem 33 .
As concentrações de DIC e seus respectivos valores de isótopos de carbono ( ) foram determinados em modo de fluxo contínuo por um analisador Aurora 1030 W TIC-TOC (OI Analytical, College Station, Texas, EUA) conectado a um espectrômetro de massa de razão isotópica ThermoFisher Delta V Plus (IRMS). A precisão de 1 − σ para medições de concentração de DIC foi melhor do que o desvio padrão relativo (dp) de 5‰. A precisão de 1 − σ para análises de foi determinada por pelo menos medições triplicadas de amostras de controle selecionadas e melhor do que ± 0,1‰. Detalhes do acoplamento do analisador OI ao IRMS e técnicas de medição são descritos em St-Jean 60 e van Geldern 61 .δδ
Amostras de POC foram filtradas em papel de filtro de fibra de vidro com poro de 0,4 μm (MN GF-5, Macherey–Nagel, Alemanha). Antes da amostragem, os papéis de filtro foram aquecidos por 6 h a 500 °C para remover traços de carbono. Após a filtração em campo, foram secos a 60 °C por 24 h e então pulverizados em um almofariz sem carbono.
Amostras pulverizadas foram secas e fumigadas com HCl concentrado em um dessecador por 24 h para remover possíveis partículas de carbonato. Este material preparado foi então pesado em cápsulas de estanho e analisado em um Analisador Elemental Costech (modelo ECS 4.010) conectado a um IRMS ThermoFisher Delta V Plus para a análise das razões de isótopos de carbono do POC (expressas como ).δ
Todos os valores de isótopos são expressos em por mil (‰) em relação ao padrão Vienna Pee Dee Belemnite de acordo com:(1)
onde R é a razão entre o isótopo de carbono pesado e o isótopo de carbono leve (ou seja, 13 C/ 12 C) 62 . Todos os dados isotópicos foram corrigidos para deriva instrumental e linearidade. Todos os desvios-padrão de 1 − σ para medições de POC de δ 13 C em pelo menos três amostras selecionadas foram melhores que ± 0,3‰.
A clorofila-a (CHL-a) foi determinada de acordo com o método de Wetzel e Likens (2000), utilizando uma solução alcalina de acetona a 90% para extração da CHL-a de filtros com poros de 0,4 μm. A absorbância foi medida em 665 e 750 nm em um espectrofotômetro para determinação das concentrações de CHL-a por litro. A CHL-a foi medida apenas em amostras selecionadas coletadas em águas superficiais (1 a 3 m).
Para aplicar o modelo de Rau et al. 55 para avaliação das relações POC, DIC e CO₂ no crescimento de algas, a pressão parcial de CO₂ ( p₂CO₂ ) precisou ser calculada. Isso foi feito conforme descrito em Marx et al. ( 2018 ), utilizando pH e HCO₂ – com a seguinte equação 63 .(2) onde HCO 3 − é a concentração de bicarbonato, H + é 10 −pH , K 1 é a primeira constante de dissociação dependente da temperatura para a dissociação de H 2 CO 3 (mol L −1 ), e K H é a constante da lei de Henry em mol L −1 atm −1 .
A composição isotópica de CO 2 ( ) foi calculada a partir do DIC e do correspondente , pH e temperatura usando constantes de equilíbrio de carbonato que foram vinculadas aos fatores de fracionamento de equilíbrio de isótopos 62 , 64 . Isso levou à seguinte equação:δδ(3) onde é o do CO 2(aq) ambiente , é o fracionamento isotópico de equilíbrio dependente da temperatura entre HCO 3 − e CO 2(aq) com a seguinte equação 64 , 65 , 66 , 67 .δδ\epsilon_{P}(4) onde b = 9,866, c = − 24,12 e T K é a temperatura em Kelvin 62 .
Esta última equação também foi confirmada por outros estudos 66 , 67 . O fator de fracionamento fotossintético entre a composição isotópica de CO 2 e POC foi calculado seguindo o modelo de Rau et al. 55 :(5) onde é a diferença de isótopos estáveis de equilíbrio dependente da temperatura entre CO 2(aq) e POC. Aqui foi derivado da Eq. ( 3 ) e é a composição de isótopos estáveis de carbono medida do POC.{\updelta }^{13} {\texto{C}}_{{{\texto{CO}}2\esquerda( {{\texto{aq}}} \direita)}} {\updelta }^{13} {\texto{C}}_{{{\texto{POC}}}}δδ
Referências
- Tanentzap, AJ et al. Construindo uma ponte entre sacos de lixo e experimentos com ecossistemas completos: uma nova abordagem para o estudo de sedimentos lacustres. J. Limnol. 76 , 431–437 (2017). ↩
- Mendonça, R. et al. Enterro de carbono orgânico em lagos e reservatórios globais. Nat. Comum. 8 , 1–7 (2017). ↩
- Raymond, PA et al. Emissões globais de dióxido de carbono de águas interiores. Nature 503 , 355–359 (2013). ↩
- Tranvik, LJ et al. Lagos e reservatórios como reguladores do ciclo do carbono e do clima. Limnol. Oceanogr. 54 , 2298–2314 (2009). ↩
- Serra-Pompei, C., Hagstrom, GI, Visser, AW & Andersen, KH. A limitação de recursos determina a resposta à temperatura das comunidades de plâncton unicelular. Limnol. Oceanogr. 64 , 1627–1640 (2019). ↩
- Webb, JR, Santos, IR, Maher, DT e Finlay, K. A importância dos fluxos de carbono aquático nos orçamentos líquidos de carbono dos ecossistemas: uma revisão em escala de bacia hidrográfica. Ecosystems 22 , 508–527 (2019). ↩
- Hanson, PC, Pace, ML, Carpenter, SR, Cole, JJ e Stanley, EH Integrando o ciclo de carbono da paisagem: necessidades de pesquisa para resolver os orçamentos de carbono orgânico dos lagos. Ecosystems 18 , 363–375 (2015). ↩
- Stets, EG, Striegl, RG, Aiken, GR, Rosenberry, DO e Winter, TC Suporte hidrológico do fluxo de dióxido de carbono revelado por balanços de carbono de todo o lago. J. Geophys. Res. 114 , G01008 (2009). ↩
- Ho, DT et al. Influência da velocidade da corrente e da velocidade do vento nas trocas gasosas entre o ar e a água em um estuário de manguezal. Geophys. Res. Lett. 43 , 3813–3821 (2016). ↩
- Read, JS et al. Dependência do cisalhamento do vento e da convecção em relação ao tamanho do lago como controles das trocas gasosas. Geofísica. Res. Lett. 39 , L09405 (2012). ↩
- Kokic, J., Wallin, MB, Chmiel, HE, Denfeld, BA e Sobek, S. A evasão de dióxido de carbono de sistemas de nascentes contribui fortemente para a exportação total de carbono de uma pequena bacia hidrográfica de um lago boreal. 120 , 16 (2015). ↩
- Lapierre, JF, Seekell, DA e Giorgio, PA Influência do clima e da paisagem em indicadores do ciclo de carbono em lagos por meio de padrões espaciais em carbono orgânico dissolvido. Glob. Change Biol. 21 , 4425–4435 (2015). ↩
- Chiu, CY et al. Cargas terrestres de matéria orgânica dissolvida impulsionam o fluxo interanual de carbono em lagos subtropicais durante períodos de seca. Sci. Total Environ. 717 , 137052 (2020). ↩
- Cole, JJ & Caraco, NF Carbono em bacias hidrográficas: conectando as perdas de carbono terrestre com o metabolismo aquático. Mar. Freshw. Res. 52 , 101–110 (2001). ↩
- Gudasz, C., Bastviken, D., Premke, K., Steger, K. & Tranvik, LJ Processamento microbiano restrito de carbono orgânico alóctone em sedimentos de lagos boreais. Limnol. Oceanogr. 57 , 163–175 (2012). ↩
- Huang, C. et al. Aumento da mineralização de carbono orgânico sedimentar induzida pelo excesso de carbono do fitoplâncton em um lago de planalto eutrófico. J. Soils Sedim. 19 (2019). ↩
- Kanduč, T., Szramek, K., Ogrinc, N. & Walter, LM Origem e ciclagem do carbono inorgânico fluvial na bacia hidrográfica do rio Sava (Eslovênia), inferido a partir dos principais solutos e isótopos estáveis de carbono. Biogeochemistry 86 , 137–154 (2007). ↩
- Savoye, N. et al. Dinâmica da matéria orgânica particulada d 15 N e d 13 C durante a floração de fitoplâncton na primavera em um ecossistema de macromarés (Baía do Sena, França). Mar. Ecol. Prog. Ser. 255 , 27–41 (2003). ↩
- Qin, B., Hu, W., Gao, G., Luo, L. & Zhang, J. Dinâmica da ressuspensão de sedimentos e o esquema conceitual de liberação de nutrientes no grande e raso Lago Taihu, China. Chin. Sci. Bull. 49 , 54–64 (2004). ↩
- Niemistö, J., Tamminen, P., Ekholm, P. & Horppila, J. Ressuspensão de sedimentos: resgate ou queda de um lago eutrófico termicamente estratificado?. Hidrobiologia 686 , 267–276 (2012). ↩
- Chen, D., Liu, Q., Xu, J. e Wang, K. Avaliação baseada em modelos do impacto de barragens hidrelétricas nas variabilidades sazonais do POC no oceano costeiro: um estudo de caso do projeto Three Gorges. J. Mar. Sci. Eng. 7 , 320 (2019). ↩
- Marcé, R. et al. Intemperismo carbonático como fator determinante da supersaturação de CO2 em lagos. Nat. Geosci. 8 , 107 (2015). ↩
- Quadra, GR, Sobek, S., Paranaíba, JR & Isidorova, A. Alto soterramento de carbono orgânico, mas alto potencial para ebulição de metano nos sedimentos de um reservatório hidrelétrico amazônico. Biogeosciences 17 , 1495–1505 (2020). ↩
- Guillemette, F., von Wachenfeldt, E., Kothawala, DN, Bastviken, D. & Tranvik, LJ Sequestro preferencial de matéria orgânica terrestre em sedimentos de lagos boreais. J. Geophys. Res. Biogeosci. 122 , 863–874 (2017). ↩
- Maavara, T. et al. Impactos de barragens fluviais no ciclo biogeoquímico. Nat. Rev. Earth Environ. 1 , 103–116 (2020). ↩
- Bravo, C., Millo, C., Covelli, S., Contin, M. & De Nobili, M. Continuum terrestre-marinho de matéria orgânica natural sedimentar em um sistema estuarino de latitude média. J. Soils Sedim. 20 , 1074–1086 (2020). ↩
- Klump, JV, Edgington, DN, Granina, L. & Remsen, CC Estimativas da remineralização e soterramento de carbono orgânico em sedimentos do Lago Baikal. J. Great Lakes Res. 46 , 102–114 (2020). ↩
- Finlay, JC & Kendall, C. em Isótopos estáveis em ecologia e ciência ambiental 283–333 (2008). ↩
- Reitsema, RE, Meire, P. & Schoelynck, J. O futuro das macrófitas de água doce em um mundo em transformação: quantidade e qualidade do carbono orgânico dissolvido e suas interações com macrófitas. Front. Plant Sci. 9 , 629 (2018). ↩
- Attermeyer, K. et al. Processamento de carbono orgânico durante o transporte através de águas interiores boreais: partículas como sítios importantes. J. Geophys. Res. Biogeosci. 123 , 2412–2428 (2018). ↩
- Bayer, TK, Gustafsson, E., Brakebusch, M. & Beer, C. As futuras emissões de carbono de lagos boreais e de permafrost são sensíveis às cargas de carbono orgânico da bacia hidrográfica. J. Geophys. Res. Biogeosci. 124 , 1827–1848 (2019). ↩
- Du, H. et al. Avaliação da eutrofização em lagos de água doce: uma nova abordagem estatística de desequilíbrio. Eco. 102 , 686–692 (2019) . ↩
- Barth, JAC, Mader, M., Nenning, F., van Geldern, R. & Friese, K. Balanços de massa de isótopos estáveis versus diferenças de concentração de carbono inorgânico dissolvido — implicações para o rastreamento da renovação do carbono em reservatórios. Isot. Environ. Health Stud. 53 , 413–426 (2017). ↩
- Chen, J. et al. Uso combinado de radiocarbono e isótopo de carbono estável para restringir as fontes e a ciclagem de carbono orgânico particulado em um grande lago de água doce, China. 625 , 27–38 (2018). ↩
- Cole, JJ et al. Encanando o ciclo global do carbono: integrando águas interiores ao balanço de carbono terrestre. Ecosystems 10 , 172–185. https://doi.org/10.1007/s10021-006-9013-8 (2007). ↩
- Tranvik, LJ, Cole, JJ e Prairie, YT. O estudo do carbono em águas interiores — de ecossistemas isolados a atores no ciclo global do carbono. Limnol. Oceanogr. Lett. 3 , 41–48 (2018). ↩
- van Breugel, Y., Schouten, S., Paetzel, M., Nordeide, R. & Sinninghe Damsté, JS O impacto da reciclagem de carbono orgânico na composição isotópica estável de carbono inorgânico dissolvido em um sistema marinho estratificado (fiorde de Kyllaren, Noruega). Org. Geochem. 36 , 1163–1173 (2005). ↩
- CETESB, CADEDSP Relação de Áreas Contaminadas , 2013). ↩
- Beghelli, FGS, Frascareli, D., Pompêo, MLM & Moschini-Carlos, V. Evolução do estado trófico ao longo de 15 anos em um reservatório tropical com baixas concentrações de nitrogênio e predominância de cianobactérias. Água, Ar, Solo, Poluição. 227 , 95 (2016). ↩
- Beghelli, FGS, Santos, ACA, Urso-Guimarães, MV & Calijuri, MC Heterogeneidade espacial e temporal em um reservatório subtropical e seus efeitos sobre a comunidade de macroinvertebrados bentônicos. Acta Limnol. Sutiãs. 26 , 306–317 (2014). ↩
- Frascareli, D. et al. Distribuição espacial, biodisponibilidade e toxicidade de metais em sedimentos superficiais de reservatórios tropicais, Brasil. Environ. Monit. Assess. 190 , 199 (2018). ↩
- Melo, DS et al. Mapas auto-organizáveis para avaliação de processos biogeoquímicos e variações temporais na qualidade da água de reservatórios subtropicais. Water Resour. Res. 55 , 14p (2019). ↩
- Simonetti, VC, Silva, DCC & Rosa, AH Análise da influência das atividades antrópicas sobre a qualidade da água da APA Itupararanga (SP), Brasil. Geosul 34 , 1–27 (2019). ↩
- Barth, JAC, Veizer, J. & Mayer, B. Origem do carbono orgânico particulado no Alto Rio Lawrence: restrições isotópicas. Planeta Terra. Ciência. Lett. 162 , 112–121 (1998). ↩
- Wei, H., Sun, J., Moll, A. & Zhao, L. Dinâmica do fitoplâncton no Mar de Bohai — observações e modelagem. J. Mar. Syst. 44 , 233–251 (2004). ↩
- Zhao, C., Zang, J., Liu, J., Sun, T. & Ran, X. Distribuição e balanço de nitrogênio e fósforo e sua influência no ecossistema do Mar de Bohai e do Mar Amarelo, China. 36 , 2115–2127 (2016) . ↩
- Suzuki, KW, Ueda, H., Nakayama, K. & Tanaka, M. Dinâmica espaço-temporal das razões de isótopos estáveis de carbono em dois copépodes oligohalinos simpátricos em relação ao máximo de turbidez estuarina (Rio Chikugo, Japão): implicações para fontes de alimento. J. Plankton Res. 36 , 461–474 (2014). ↩
- Topçuoğlu, BD, Meydan, C., Nguyenc, TB, Langc, SQ e Holdena, JF Cinética de crescimento, fracionamento de isótopos de carbono e expressão gênica no hipertermófilo Methanocaldococcus jannaschii durante crescimento limitado por hidrogênio e transferência de hidrogênio entre espécies. Appl. Environ. Microbiol. 18 , 00180–00181 (2019). ↩
- Português Ravelo, AC & Hillaire-Marcel, C. em Proxies em desenvolvimentos paleoceanográficos do Cenozóico tardio em geologia marinha (eds C. Hillaire–Marcel & A. De Vernal) 735–764 (Elsevier, 2007). ↩
- Striegl, RG, Aiken, GR, Dornblaser, MM, Raymond, P. & Wickland, KP Uma diminuição na exportação de COD normalizada pela vazão pelo Rio Yukon durante o verão e o outono. Geofísica. Res. Lett. L21413 (2005). ↩
- Yoshioka, T. Fracionamento de isótopos de carbono fitoplanctônico: equações que explicam os mecanismos de concentração de CO₂ . J. Plankton Res. 19 , 1455–1476 (1997). ↩
- Bottino, F., Calijuri, MC & Murphy, KJ Variação temporal e espacial de variáveis limnológicas e biomassa de diferentes espécies de macrófitas em um reservatório neotropical (São Paulo — Brasil). Acta Limnol. Bras. 25 , 387–397 (2013).CAS Google Acadêmico ↩
- Taniwaki, RH et al. Estrutura e dinâmica da comunidade de algas perifíticas em um reservatório subtropical (estado de São Paulo, Brasil). Acta Botânica Brasilica 27 , 551–559 (2013). ↩
- Baird, ME, Emsley, SM e McGlade, JM: Usando um modelo de crescimento do fitoplâncton para prever o fracionamento de isótopos estáveis de carbono. J. Plankton Res. 23 , 841–848 (2001). ↩
- Rau, G.H.; Riebesell, U.; Wolf-Gladrow, D. (1996). Um modelo de fracionamento fotossintético de 13C pelo fitoplâncton marinho baseado na captação molecular difusiva de CO₂. Mar. Ecol. Prog. Ser. 133, 275–285. ↩
- Kottek, M., Grieser, J., Beck, C., Rudolf, B. & Rubel, F. Mapa-múndi da classificação climática de Köppen-Geiger atualizado. Meteorol. Z. 15 , 259–263 (2006). ↩
- Pedrazzi, FJDM, Conceição, FTD, Sardinha, DDS, Moschini-Carlos, V. & Pompêo, M. Qualidade espacial e temporal da água no reservatório de Itupararanga, Bacia do Alto Sorocaba (SP), Brasil. J. Recursos Hídricos. Prot. 5 , 64–71 (2013). ↩
- Ribeiro, AR, Biagioni, RC & Smith, WS Estudo da dieta natural da ictiofauna de um reservatório centenário. Iheringia 104 , 404–412 (2014). ↩
- Smith, WS & Petrere, M. Jr. Padrões espaciais e temporais e sua influência na comunidade de peixes do reservatório de Itupararanga, Brasil. Rev. Biol. Trop. 56 , 2005–2020 (2008). ↩
- St-Jean, G. Análise quantitativa e isotópica automatizada ( 13 C) de carbono inorgânico dissolvido e carbono orgânico dissolvido em fluxo contínuo usando um analisador de carbono orgânico total. Rapid Commun. Mass Spectrom. 17 , 419–428 (2003). ↩
- van Geldern, R. et al. Análise de isótopos estáveis de carbono de carbono inorgânico dissolvido (CID) e carbono orgânico dissolvido (COD) em águas naturais — resultados de um teste de proficiência mundial. Rapid Commun. Mass Spectrom. 27 , 2099–2107 (2013). ↩
- Clark, ID & Fritz, P. Isótopos ambientais em hidrogeologia (CRC Press, Londres, 1997). ↩
- Marx, A. et al. Dados de águas subterrâneas melhoram a modelagem da liberação de CO₂ de córregos de cabeceira com uma abordagem de isótopos DIC estáveis. Biogeosciences 15 , 3093–3106 (2018). ↩
- Mook, WG, Bommerson, JC & Staverman, WH Fracionamento de isótopos de carbono entre bicarbonato dissolvido e dióxido de carbono gasoso. Planeta Terra. Sci. Lett. 22 , 169–176 (1974). ↩
- Mayer, B. et al. Avaliação da utilidade da composição isotópica de CO₂ para monitoramento de vazamentos em locais de armazenamento de CO₂ : uma revisão. Int. J. Greenhouse Gas Control 37 , 46–60 (2015). ↩
- Myrttinen, A., Becker, V. & Barth, JAC. Retificação para ‘Uma revisão dos métodos utilizados para investigações de fracionamento de isótopos de equilíbrio entre carbono inorgânico dissolvido e CO₂’ [Earth Sci. Rev. 115 (2012) [192–199]]. Earth-Sci. Rev. 141 , 178 (2015). ↩
- Myrttinen, A., Becker, V. & Barth, JAC Uma revisão de métodos usados para investigações de fracionamento de isótopos de equilíbrio entre carbono inorgânico dissolvido e CO 2 . Earth Sci. Rev. 115 , 192–199 (2012). ↩